
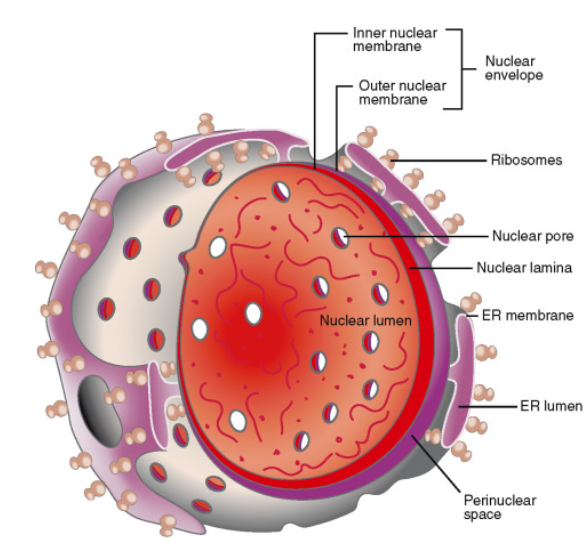
The nuclear membrane is also known as the nuclear envelope or perinuclear cisternae. It encloses the nuclear region containing the DNA and defines the nuclear compartment.
Structure of Nuclear Envelope
- The nuclear membrane is formed of two concentric unit membranes.- the inner and outer membrane.
- The space between the inner and outer membrane is called perinuclear space.
- The nuclear membrane is not a complete circle and has small openings in between which are called nuclear pores.
- In addition, the nuclear lamina, a dense protein network made of intermediate filaments provides mechanical support to the membrane.
Inner Nucelar Membrane
- The inner nuclear membrane consists of specific proteins that are binding sites for supporting the sheath of intermediate filaments, nuclear lamina, etc.
- The nuclear lamina is in contact with chromatin and nuclear RNAs.
Outer Nucelar Membrane
- The outer nuclear membrane surrounds the inner membrane and is continuous with the endoplasmic reticulum.
- It resembles the membrane of the endoplasmic reticulum.
- It is often surrounded by intermediate filaments on the outside.
- The outer surface of the nuclear envelope is studded with ribosomes for protein synthesis.
- Proteins made by these ribosomes are transported into the perinuclear space.
Perinuclear Space
The perinuclear space is filled with the fluid that is continuous with the endoplasmic reticulum lumen. This fluid may contain crystalline deposits, fibers, electron-dense material, lipid droplets, etc.
Nuclear Lamina
Nucelar lamina is also called fibrous lamina, internal dense lamina, zonula nucleum limitans, nucelar cortex, and lamina densa.
- It is a protein network of 50-80 nm thick.
- It lines the inner surface of the inner membrane except at nuclear pores
- Nucelar lamin has a square lattice of intermediate filaments.
- These lamins form dimers with rod-like domains and two globular heads.
- When the pH is favorable, the dimers spontaneously associate with intermediate filaments.
- It has a dynamic structure.
- In mammals, the dimers disassemble into tetramers of hypophosphorylated lamin A and lamin C and membrane-associated lamin B during mitosis.
- This causes lamin A and C to become soluble and dephosphorylated during telophase.
- Lamins are crucial for the assembly of interphase nuclei.
Nuclear Pore
The nuclear membrane is not altogether continuous but is broken by nuclear pores or openings at several places. Nuclear pores are small openings or perforations seen in the nuclear envelope. These pores appear circular on the surface view, with a diameter of 10-100nm. The molecular weight of each nuclear pore complex is 50-100 m daltons.
Although it was believed that a nuclear pore is similar to a diaphragm, it was later found that it has a more complex structure often called the nuclear pore complex.
Nuclear Pore Complex Structure
Nuclear pores are enclosed by a circular structure called annuli. The pores and annuli together form the pore complex.
- It has an octagonal symmetry.
- Annular material is an amorphous cylinder that extends through the pore into either side of the pore margin.
- It has an outer diameter of about 120°A which exceeds that of pore.
- The central pole of the annulus varies from 300-500A.
- The annular material is protein.
- Each pore complex has two parts- an inner ring and an annuli having a diameter of 80 nm and a larger central plug.
- Radial spokes arising from the central plug connect it to the rings.
- Rings have particle P attached to them and they are inactive ribosomes.
- The center of the pore complex has a hole containing aqueous channel.
- This channel facilitates the transportation of water-soluble molecules between the cytoplasm and the nucleus.
- Each pore has the lipid bilayer of the outer and inner nuclear membrane connected but stays chemically distinct.
- A glycoprotein anchors a covalently bound acetyl neuraminic acid seen on the cytoplasmic side excluding the nuclear pore.
- The second protein helps with transcription and materials through the nuclear pore.
- The number of pores varies with the species and this number will be approximately 5-36% of the surface area of the nuclear membrane.
- Pore density is related to the transcriptional activity of cells.
- Nuclear pores are evenly or randomly arranged in somatic cells.
- It may be in rows in spores, clusters in oocytes, and hexagonal in Malphigian tubules.
Nuclear Pore Functions
- Nuclear membrane pores are the pathways for the exchange of macromolecules.
- The annuli regulate their exchange through the pore complex according to their chemical nature and size.
- The differential properties of the cell are dependent on the nature of the annuli.
Nucelo-Cytoplasmic Traffic
The nuclear pore complex helps with transportation between the nucleus and cytoplasm. The main function of the nuclear envelope is to prevent ribosomes from entering the nucleus.
So, the nuclear pore facilitates the movement of only ions, regulatory and structural proteins, nucleotides, and catalysis proteins from the cytoplasm to the nucleus. mRNA, tRNA, and other subunits of ribosomes are transported from the nucleus to cytosol.
- The opening of the pore is activated by signals on large nuclear proteins having nuclear import signals or karyophillic signals.
- A nuclear protein called nuceloplasmin has distinct head and tail domains. The tail domain consists of a signal that enables its folding and transportation through the membrane.
- Each nuclear pore transports about 100 histone proteins into the nucleus per minute.
- An actively growing cell has three newly assembled ribosomes exported per minute from the nucleus to the cytosol.
- The ribosomal export is an active transportation through the nuclear pore.
References
- Agarwal, P. V. |. V. (2004). Cell biology, Genetics, Molecular Biology, Evolution, and Ecology: Evolution and Ecology. S. Chand Publishing.
- (2007). Regulation of Gene Expression. Medical Cell Biology (Third Edition), 149-190. https://doi.org/10.1016/B978-0-12-370458-0.50010-4
Additional Reading
- Structure and Function of Golgi Apparatus
- Types of Plastids and Their Functions
- Ribosomes: Structure and Function
- Endoplasmic Reticulum in Plant Cell



